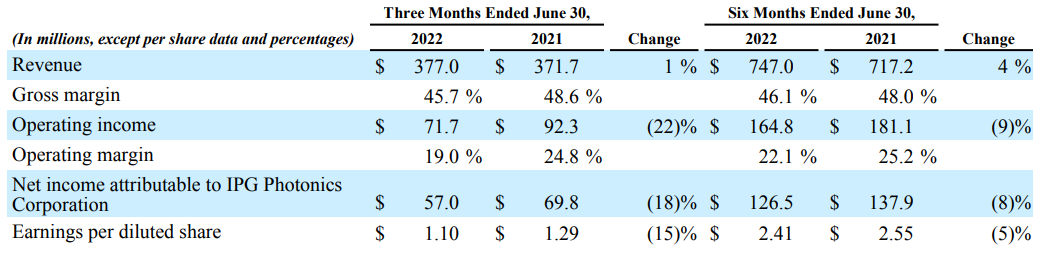
肿瘤免疫微环境的异质性
前言
在肿瘤发生和随后的转移过程中,恶性细胞逐渐多样化,变得更具异质性。因此,肿瘤可能被多种免疫相关成分浸润,包括细胞因子/趋化因子环境、细胞毒性活性或免疫抑制因子。这种免疫异质性在几乎所有实体瘤中普遍存在,并且随着肿瘤的发展以及治疗干预而在空间上或时间上发生变化。抗肿瘤免疫的异质性与疾病的进展和治疗的反应性密切相关,尤其是在免疫治疗领域。
因此,准确了解肿瘤免疫异质性对于有效治疗的发展至关重要。在多区域和组学测序、单细胞测序以及纵向液体活检方法的帮助下,最近的研究显示了研究肿瘤免疫异质性的复杂性及其在免疫治疗中临床相关性的潜力。探索肿瘤免疫微环境异质性的机制,有助于我们对肿瘤异质性的临床评估,从而促进更有效的个性化治疗的发展。
肿瘤免疫微环境异质性的起源

遗传不稳定性
高通量测序方法长期以来一直被用来描述肿瘤细胞的突变谱和进化轨迹,这些研究在时空维度上描绘了广泛的遗传肿瘤异质性,包括异质性单核苷酸突变、插入、缺失和拷贝数突变。在肿瘤进展过程中,遗传不稳定性导致随机产生这些改变。
在原发性肿瘤中,受驱动基因的突变通常会带来生存优势;因此,这些细胞更有可能占据生长优势的位置,并发展成为优势克隆群体。相比之下,乘客突变在肿瘤进化过程中不会带来显著的生长优势,它们被认为是亚克隆肿瘤细胞的主要来源。因此,起源于克隆和亚克隆肿瘤细胞的遗传不稳定性构成了肿瘤进化和时空异质性的基础。同时,这种基因异质性塑造了肿瘤的抗原谱,并最终促成了肿瘤免疫微环境的异质性。
表观遗传修饰
越来越多的证据表明,肿瘤细胞的表观遗传重塑也参与了异质性肿瘤免疫微环境的形成。这种调节机制主要归因于DNA修饰的改变、染色质可及性的改变或转录后水平上的基因表达调节,如非编码RNA干扰。这些表观遗传修饰促进了肿瘤细胞的恶性进展,并有助于形成肿瘤免疫微环境。
除了甲基化之外,各种染色质和表观遗传重塑机制赋予肿瘤细胞适应周围环境的优势。通常,表观遗传修饰是有条件可逆的。在肿瘤细胞中,这些修饰可以由其后代遗传,因此,这些细胞在空间和纵向维度上表现出显著的异质性。
微环境扰动适应度
肿瘤细胞持续暴露于细胞外微环境扰动中。越来越多的证据表明,细胞内适应性可由外部应激引起,包括DNA损伤反应、未折叠蛋白质反应和线粒体应激信号。肿瘤在组织学和血管构筑方面表现出显著的异质性。
肿瘤内血管近端或远端的区域可能暴露于不同的氧气供应。因此,免疫成分能够以时空异质的方式适应基于氧气张力、葡萄糖可用性或氧化途径的外部刺激。无论免疫组分在缺氧条件下是否适应良好,几乎所有的缺氧反应都与肿瘤免疫微环境的重编程密切相关,其主要特征是细胞糖酵解代谢的局部转换、葡萄糖消耗增加、丙酮酸和乳酸产生增加,和酸化。
抗肿瘤治疗的反应
在治疗过程中,微环境中的肿瘤细胞和所有免疫成分要么被打击(如放疗),要么持续暴露于抗肿瘤药物。为了应对这些应激因素,肿瘤和免疫细胞的适应性机制被启动,以建立新的内环境平衡。
由于驱动突变或分子特征的内在异质性,肿瘤细胞对治疗的反应性存在显著差异。细胞毒性条件使肿瘤和免疫细胞经历表型改变、细胞衰老甚至细胞死亡。无法在治疗中存活的局部肿瘤克隆通过自噬介导的细胞死亡释放大量ATP。这些ATP可以促进趋化作用,并在肿瘤中产生炎症反应。相反,在存在细胞外核苷酸酶的情况下,ATPs可以在细胞外基质中迅速转化为腺苷,从而形成抑制性免疫微环境。
对于免疫细胞而言,T细胞表型在对ICI的反应中发生显著变化,伴随着不同的T细胞亚群组成和细胞因子产生。治疗药物、肿瘤细胞和免疫细胞之间复杂而动态的相互作用显著促进了时空异质免疫微环境的形成。
肿瘤免疫微环境的异质性
空间异质性
肿瘤免疫微环境的特征主要由肿瘤和非肿瘤成分决定。它们的定位或丰度/活性在空间上是不同的,包括抑制性免疫检查点(如PD-L1)的表面表达、免疫抑制或促炎细胞因子的分泌、免疫抑制或效应细胞的浸润、血管系统的状态、边缘区域的空间距离,以及代谢营养素的分布。这些空间变异也会对临床预后和治疗反应产生深远影响。

肿瘤内T细胞的表型表现出显著的异质性。T细胞通常具有不同的克隆性、增殖潜能、分化阶段、功能极化、细胞因子分泌谱或代谢环境。关于T细胞库的倾向性,扩增/增殖性T细胞受体(TCR)可进一步分为常见TCR克隆(在肿瘤内所有区域检测到)或区域克隆(异质分布)。常见和区域性TCR克隆的数量与常见和区域性非同义突变的负荷呈正相关,表明区域异质性、抗原驱动的T细胞增殖。此外,值得注意的是,调节性T细胞(Treg)也显示出显著的肿瘤内空间异质性和功能取向。
除了T细胞亚群,在各种肿瘤类型中也发现了许多其它免疫细胞的肿瘤内异质性。在胃癌中,CD68+CD163+CD206+表型的巨噬细胞主要位于基质中,与边缘区相比,CD68+IRF8+巨噬细胞在核心区过表达。除了免疫细胞群,基质细胞(如成纤维细胞)在肿瘤中也表现出高度的空间取向性。
代谢谱是免疫微环境的重要调节因子,可能通过影响癌细胞的增殖潜能和适应环境而发挥作用。代谢特征的异质性似乎有助于肿瘤免疫微环境的异质性。具有高糖酵解活性的恶性细胞不仅可以将其代谢途径转变为合成代谢反应,还可以产生大量免疫抑制介质,如乳酸和腺苷,以减弱细胞毒性细胞的免疫监视。
时间异质性
肿瘤和免疫细胞很容易被遗传或非遗传环境因素干扰,因此决定了疾病的进展和对抗肿瘤治疗的反应,以及肿瘤细胞本身的动态进化。在疾病从非侵袭性病变发展为侵袭性表型的过程中,胰腺导管腺癌患者的RNA-Seq揭示了免疫细胞浸润成分的显著变化。通常表现为CD8+T细胞和树突状细胞的浸润减少,以及免疫抑制细胞(包括Treg、MDSC或CAF)的异常聚集或扩增。
此外,多种肿瘤类型的疾病进展过程中,还存在细胞溶解活性受损、细胞库扩展和克隆性受限以及T细胞和B细胞的进行性衰竭。个体患者中免疫不利区域或病变的出现似乎与疾病控制和生存预后成反比,这进一步加强了时空异质性对疾病结果的重要性。
肿瘤免疫微环境异质性的临床意义
几乎所有癌症患者的免疫预后或预测性生物标志物都是基于对单个活检样本的分析而建立的。然而,异质性是研究重现性的一个重大障碍,并削弱了其临床实用性。此外,大量证据表明,肿瘤微环境的异质性,无论是遗传的还是免疫的,都会影响实体瘤患者的免疫治疗效果。
PD-L1表达的异质性
自从第一个证据支持PD-L1蛋白表达和抗PD-1检查点阻断疗法的疗效相关以来,PD-L1水平已被用作伴随诊断,以预测各种实体瘤类型对ICI免疫疗法的临床反应。然而,无论在肿瘤内或肿瘤间尺度上,还是在空间和时间维度都存在显著的异质性PD-L1表达。
在评估来自NSCLC患者的原发性和脑转移肿瘤中PD-L1的表达后,可以观察到两种病变之间PD-L1表达存在显著差异。PD-L1表达是由IFN-γ信号通路强烈诱导的,该信号通路在含有功能异常JAK1/2突变的亚克隆中受到异质性调控。此外,在接受ICI治疗的患者中出现的抗原处理和呈递缺陷亚克隆与黑色素瘤、肺癌和结直肠癌的不良临床结果相关。这种潜在的异质性可以解释为什么一部分PD-L1阳性肿瘤患者没有反应,而一些PD-L1阴性肿瘤患者对ICI免疫疗法反应良好。
TMB高反应患者的异质性反应
突变负荷是新抗原负荷的一种合理近似替代物,已被用于在各种实体瘤类型中识别ICI免疫疗法的有利应答者。然而,TMB高的患者对ICB治疗的反应是高度异质性的,相当一部分TMB水平较低的患者也可以从ICI免疫疗法中获益,反之亦然。
对于对ICI治疗反应不佳的TMB高风险患者,缺陷/失调的抗原呈递机制被认为是免疫治疗抵抗的主要机制,尤其是HLA的单倍型和区域表达,以及B2M分子的表达。此外,系统发育分析发现,克隆异质性,无论是通过构成肿瘤的克隆数量来衡量,还是通过克隆分化来衡量,都对ICI治疗的生存结果产生了深远的影响。
MMR缺陷患者的异质性反应
缺乏MMR的患者对ICI治疗极为敏感,这在很大程度上归因于升高的预测新抗原和改善的免疫原性肿瘤微环境。在众多实体瘤类型的晚期dMMR患者中,pembrolizumab作为MSI-H肿瘤患者的最佳治疗方法获得了前所未有的认可。然而,只有少数患者对ICI治疗反应良好。
dMMR肿瘤的肿瘤细胞内在基因型和外在免疫环境都可以影响其疗效,在dMMR肿瘤中,浸润免疫细胞基因组不稳定性的程度似乎在很大程度上是异质性的,导致免疫原性有限的离散生态位和免疫介导的肿瘤控制不足,这可能导致耐药性。此外,影响MSI-H肿瘤对ICI反应的其他变量包括,在发育过程中,它们倾向于执行强大的免疫编辑和转化为糖酵解谱,这在很大程度上有助于免疫逃逸。
克服肿瘤免疫微环境异质性的策略

针对公共抗原
存在空间上不同的免疫原性是免疫治疗产生异质性反应的根本原因。克服这一障碍的合理策略包括开发可针对整个肿瘤生态位的共享新抗原或均匀表达的肿瘤相关抗原(TAA)。
与个性化的新抗原相比,公共或共享的新抗原来源于癌基因的驱动突变或基因组中的其他热点突变。它们的特征是具有特定癌症亚型的患者子集中呈现的免疫原性表位。因此,公共新抗原的发现依赖于从相当大的患者库中分析个体化新抗原。公共新抗原的一个例子是KRAS上G12D的突变,该突变在胰腺癌、结肠腺癌、非小细胞肺和结直肠癌中常见。类似地,TP53是一种众所周知的肿瘤抑制基因,在大量癌症中广泛突变,具有广泛的热点突变,并为多种癌症所共有。
多抗原靶向
通过同时靶向多个抗原来克服免疫原性异质性是另一个合理的途径。这种策略最大限度地防止实体瘤中由于异质性免疫原性而导致抗原丢失时免疫逃逸的可能性。
目前,多种双靶点策略已经应用于CAR-T细胞治疗,两种分别靶向不同表位的CAR-T细胞,或通过同时靶向多个表位的单个CAR-T细胞。例如,抗CD38/BCMA CAR-T细胞已在多发性骨髓瘤患者,抗CD19/CD22双特异性CAR-T细胞在治疗B-NHL患者中进行了测试。
促进免疫原性细胞死亡和表位扩散
在癌症免疫治疗中,肿瘤疫苗是一种很有前途的治疗策略,不仅因为它能够通过传递高度免疫原性抗原来引发炎症环境,还因为它们通过促进表位扩散来拓宽和多样化抗原谱。
随着个人新抗原功能鉴定的突破,在长期临床过程中,使用合成新抗原治疗的患者不断检测到强大的新抗原特异性T细胞反应。随着新抗原特异性T细胞克隆的持续存在,疫苗接种后还观察到具有更广泛规格的多样化T细胞库,从而提供了更大的可能性,以异种抗原性完全覆盖肿瘤块。除了直接提供合成肽外,编码个性化新抗原的RNA疫苗或负载树突状细胞的新抗原疫苗也成功地动员了具有广泛T细胞特异性的有效和持续的抗肿瘤免疫。
克服免疫微环境异质性的另一种策略是利用溶瘤病毒直接杀死恶性细胞,通过释放大量免疫活性成分,包括隐匿的肿瘤相关抗原、危险信号、细胞因子和趋化因子,强烈促进免疫原性细胞死亡。这种伴随的旁分泌作用对于激活肿瘤内空间周围位点的非选择性T细胞毒性至关重要,尤其是对于免疫原性相对较低的位点。除溶瘤病毒外,还可以引入各种方法,通过可溶性成分介导的机制激发炎症免疫微环境,例如新型药物、酪氨酸激酶抑制剂、阳离子两性肽、微波、放射治疗。所有这些都强调了一个关键问题,即强有力的治疗反应通常涉及整个免疫微环境向免疫活性、同质环境的重塑。
小结
肿瘤发生是一系列遗传和非遗传过程的综合和累积失调。由于肿瘤基因组固有的遗传不稳定性,在疾病进展过程中,大部分致瘤事件不可避免地以随机方式发生。这些随机事件在空间或时间维度上为异质性免疫微环境的发展创造了必要的条件。此外,对代谢物和营养素的竞争、治疗压力或关键癌基因的进化不断重塑免疫微环境。这最终为恶性细胞逃避免疫监视创造了机会,最终导致疾病进展和转移。
这种免疫异质性也是基于单个活检的预测性生物标记物表现不佳以及对免疫疗法产生耐药性的原因。因此,考虑到肿瘤基因组的不稳定性和异质性的无休止发展,有必要关注从异质性肿瘤模型中吸取的经验教训。一个复杂的、受控的模型使我们能够精确地理解对异质性的抗肿瘤免疫反应的调节机制。而治疗方法不仅应考虑致癌靶点或代表性免疫检查点,还应考虑免疫微环境的异质性和反应性。跟踪肿瘤和免疫细胞之间的时空相互作用对于指导免疫疗法的有效而持久的反应至关重要。

原文标题:肿瘤免疫微环境的异质性
郑重声明:文章仅代表原作者观点,不代表本站立场;如有侵权、违规,可直接反馈本站,我们将会作修改或删除处理。
相关阅读



猜你喜欢
-
天津市印发加快推进城镇环境基础设施建设实施方案!
2022-08-01 -
肿瘤代谢重编程与免疫应答
2022-07-19 -
肿瘤免疫治疗中的纳米技术
2022-07-11 -
艾迈斯欧司朗推出新型环境光/接近传感器
2022-07-06 -
肿瘤细菌免疫治疗的研究现状与未来方向
2022-07-05 -
8月1日起施行!财政部 生态环境部等七部门发布《中国清洁发展机制基金管理办法》
2022-07-04 -
肿瘤免疫治疗中的非凋亡调节性细胞死亡
2022-07-01 -
北京市生态环境局等七部门印发《北京市“十四五”时期低碳 试点工作方案》
2022-07-01 -
肿瘤部位的适应性免疫抵抗
2022-06-28 -
肿瘤免疫中调节性T细胞浸润的机制
2022-06-27 -
生态环境部:推进生态环保金融支持项目储备库、国家气候投融资重点项目库建设
2022-06-24 -
半导体车间背后的环境问题
2022-06-23 -
7部门印发方案推进减污降碳协同增效
2022-06-21